What Loneliness Actually Does to the Brain
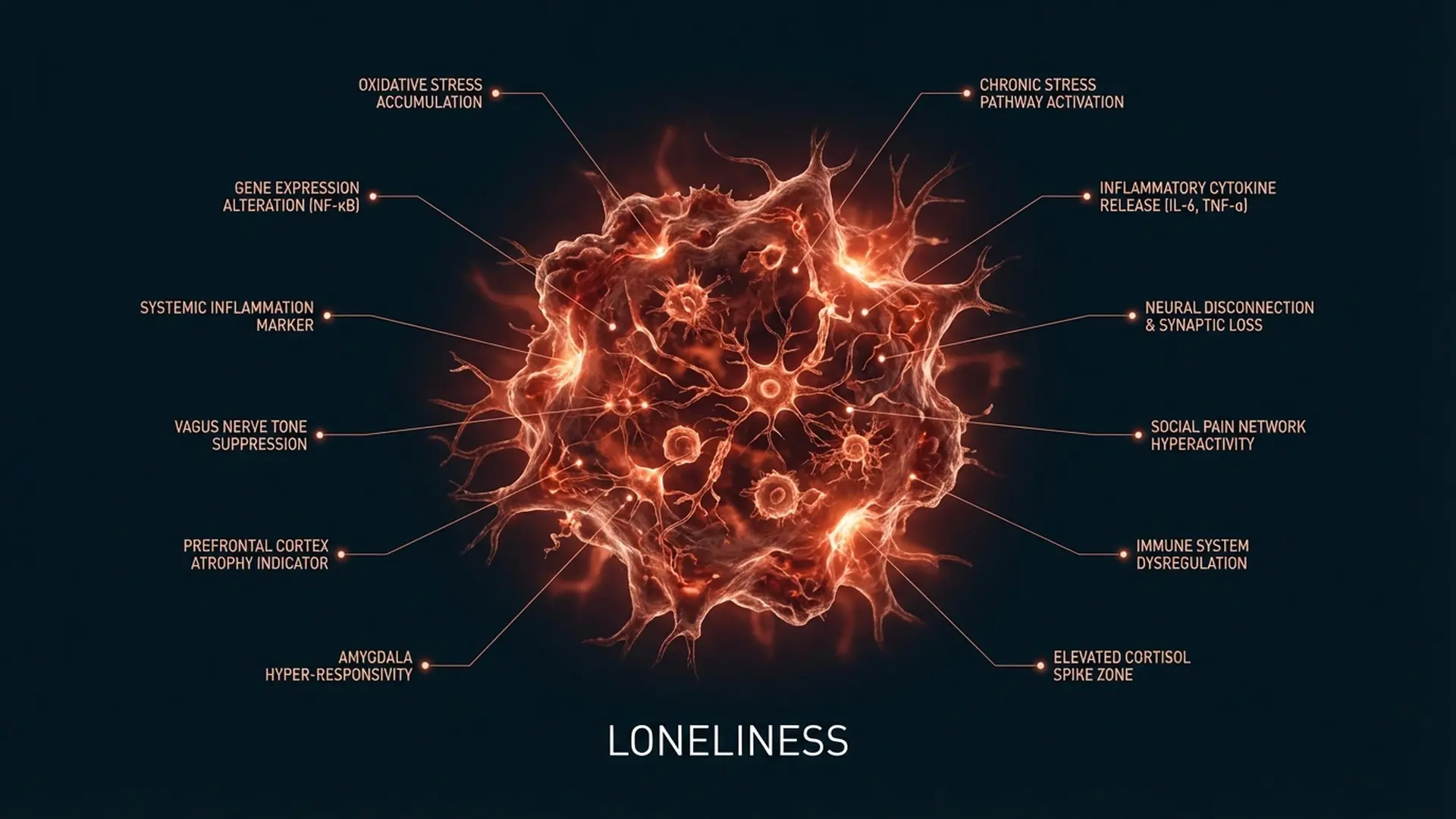
Loneliness is not a feeling. It is a neurological state: one that triggers measurable inflammatory cascades, disrupts cortisol regulation, and activates the brain’s threat-detection circuitry in precisely the same way physical danger does. The brain does not distinguish between social exclusion and physical harm. Both route through the same alarm system. Both produce the same physiological mobilization. This is not a metaphor. It is a documented mechanism.
When I work with high-capacity individuals who report loneliness, the first correction I make is this one: they are not suffering from a personal failing. Their brain is doing exactly what it was trained to do. Loneliness is a survival signal, not a character flaw. The problem is that in modern high-performance environments, that signal misfires chronically, and the neural and physiological consequences accumulate silently, long before the individual recognizes the damage.
In my practice, I consistently observe that this state arrives with a specific signature. It is not sadness. It is a particular kind of hypervigilance: a low-grade scanning of the environment for threats that do not materialize, a heightened sensitivity to ambiguity in social interactions, and a fatigue that sleep does not resolve. Understanding why this happens at the neural level is the first step toward intervention.
The Neuroscience of Loneliness: Inflammatory Cascades and Threat Circuitry
John Cacioppo’s research at the University of Chicago established that loneliness activates the hypothalamic-pituitary-adrenal (HPA) axis: the same circuit responsible for cortisol release during physical threat. Chronically elevated cortisol suppresses immune function, accelerates cellular aging, and degrades the prefrontal cortex’s capacity for executive function. A person experiencing sustained loneliness is, at the biochemical level, under continuous physiological stress.
The inflammatory response compounds this. Research published in the journal PNAS demonstrated that chronic social disconnection upregulates the expression of pro-inflammatory genes: specifically, those associated with the CTRA (conserved transcriptional response to adversity) profile. This is the same genomic signature observed in individuals facing social threat, predator exposure, or physical wounding. The body treats social disconnection as injury.
Neuroimaging data extends this further. The dorsal anterior cingulate cortex and anterior insula, regions that process physical pain, show elevated activation during social exclusion. Naomi Eisenberger’s work at UCLA using the “Cyberball” paradigm demonstrated that being excluded from a virtual ball-tossing game produced dorsal ACC activation indistinguishable from the response to mild physical pain. Social pain is not analogous to physical pain. At the level of neural substrate, they share the same circuitry.
This has practical consequences. Loneliness degrades the very neural resources needed to resolve it. Sustained threat-circuit activation reduces prefrontal inhibitory control, narrows attentional scope, and increases hypervigilance to social cues, making ambiguous interactions read as hostile, and neutral faces read as rejecting. The brain begins to confirm the disconnection story it is already running.
Why Social Pain and Physical Pain Share the Same Neural Address
The anterior cingulate cortex (ACC) is one of the most consequential structures in the human brain. It integrates cognitive, emotional, and autonomic information. It monitors error signals, detects social conflict, and, critically, processes the affective component of pain. Not the location of pain. The suffering of pain. The ACC is where pain becomes distressing rather than merely sensory.
When social disconnection occurs (rejection, exclusion, perceived abandonment) the ACC activates as though tissue has been damaged. This is not a malfunction. It is the evolutionary mechanism. For a species that survived through group membership, social rejection carried genuine survival risk. The brain developed pain circuitry around social loss for the same reason it developed it around injury: because both threatened survival.
What this means practically is that telling someone experiencing this state to “think positively” or “put themselves out there” is neurologically equivalent to telling someone with a broken arm to walk it off. The pain is real, it is rooted in tissue-level neural activation, and it requires intervention at the level of the nervous system, not at the level of attitude or willpower.
In my work, I encounter this misunderstanding with striking regularity. Highly intelligent, analytically capable individuals have been told, by themselves and by others, that their disconnection is a problem of perspective. That reframe fails because it addresses cognition while bypassing the subcortical circuitry that is actually driving the state. The ACC does not respond to positive thinking. It responds to neural safety. Those are different targets, and they require different interventions.
The Loneliness Paradox: Why High-Performers Experience Disconnection Differently
There is a particular form of loneliness that does not fit the standard portrait. It is not the disconnection of isolation or social poverty. It is the condition of the person who is constantly surrounded by people (colleagues, clients, family, a full calendar) and who cannot access genuine connection in any of it. This is not a failure of social skill. It is a product of neural adaptation to high-performance environments.
When I work with C-suite clients, founders, and high-capacity professionals, I see this pattern with consistency: the same neural efficiency that drives exceptional performance also accelerates social disconnection. The prefrontal cortex systems that produce strategic clarity, emotional regulation under pressure, and rapid decision-making also create a kind of cognitive distance from the social environment. The brain becomes calibrated for performance, and ordinary social interaction fails to register as meaningful or rewarding.
In this population, the state is often compounded by a secondary layer: the belief that it should not be present. “I have everything. I am surrounded by people who respect me. I have no reason to feel this way.” That sentence, some version of it, appears in my work repeatedly. It represents the gap between external circumstance and internal neural state. The ACC does not evaluate social satisfaction against one’s career achievements. It evaluates the quality of felt social connection. Those are independent measures.
High-performers also experience a specific cortisol pattern that intensifies the disconnection: the sustained cortisol elevation of chronic high-stakes performance primes the threat circuit and reduces the reward signal associated with social contact. Dopamine and oxytocin responses to connection are blunted when the stress axis is chronically activated. The capacity to feel the rewarding aspects of social engagement is neurologically suppressed. This means the condition can worsen even when social contact increases, because the neural reward system is not in a state to register that contact as meaningful.
Loneliness as a Neural Alarm Signal
The most important reframe I offer clients experiencing loneliness is this: the signal is not the problem. The uncorrected signal is the problem.
Loneliness is the brain’s alarm system for social disconnection: a core signal within the domain of social resilience and connection. Like hunger signals nutritional deficit, or pain signals tissue damage, it communicates that something the nervous system requires, genuine felt connection, is absent. The signal is appropriate. The signal is functional. In a world where social connection was proximate and reliable, the alarm would fire briefly and resolve. The problem in modern life is that the conditions that would resolve it are structurally absent or neurologically inaccessible, and the alarm runs continuously.
Cacioppo described this as the difference between transient loneliness (acute signal, resolves with connection) and chronic loneliness (signal locked open, threat circuit chronically activated). The transition from transient to chronic is not about the amount of social contact. It is about the neural state of the individual receiving that contact. A brain running chronic threat-circuit activation cannot properly metabolize social connection. The alarm stays on regardless of input.
What changes this is not more social activity. What changes this is neural recalibration: specifically, bringing the threat circuit out of chronic activation, restoring HPA axis baseline, and rebuilding the neural conditions in which genuine felt connection becomes possible. This is not achievable through behavioral strategies alone. It requires working at the level of the nervous system directly.
In my practice, I have observed that when the nervous system is recalibrated, when the ACC threat response comes down from chronic activation, individuals often describe a qualitative shift in how they experience the social interactions they were already having. The interactions were there before. The neural capacity to register them as meaningful was not. This distinction, between social contact and felt connection, is the central insight that changes the intervention approach entirely.
The Cortisol-Loneliness Loop That Locks the Pattern In
Loneliness elevates cortisol. Elevated cortisol increases threat vigilance. Increased threat vigilance reads ambiguous social signals as hostile. Hostile social interpretation increases perceived rejection. Increased perceived rejection deepens the state. This is not a metaphorical cycle. Each step in this sequence has a measurable neural correlate, and each step activates the next through documented biochemical pathways.
The practical consequence is that loneliness is self-reinforcing at the neural level, not because the person is doing anything wrong, but because the brain is doing exactly what a chronically threat-activated brain does. It protects itself by increasing vigilance and decreasing social risk-taking. The result is further withdrawal from the contact that might interrupt the cycle.
What makes this particularly difficult for high-performers is the absence of feedback from trusted others. In most social environments, a friend or partner will notice behavioral withdrawal and offer corrective signal. In high-performance professional environments, withdrawal often reads as focus, as strength, as appropriate executive composure. The social feedback loop that might interrupt the cycle is absent. The pattern runs undisturbed for years.
By the time many of my clients arrive, this pattern has been running for a decade or longer. The neural circuitry has been strengthened through repetition. The hypervigilance has become automatic. The blunting of social reward has become baseline. This is not irreversible, neuroplasticity ensures it is not, but it requires deliberate, sustained neural-level intervention to change.
Dr. Ceruto’s Social Reconnection Methodology
The standard response to loneliness (join a group, volunteer, increase social activity) addresses behavior. It does not address the neural state that determines whether behavior produces felt connection. This is why so many intelligent, motivated people try those recommendations and find them ineffective. They are applying the right solution to the wrong level of the problem.
My methodology targets the neural infrastructure first. The Real-Time Neuroplasticity™ approach to social disconnection works in three phases. The first phase is threat-circuit deactivation: identifying the specific patterns of hypervigilance and negative-interpretation bias that are keeping the ACC in chronic activation, and intervening on those patterns in real time, during the moments when they are actually running, not retrospectively in a structured session environment.
The second phase rebuilds the neural conditions for reward-responsiveness to social contact. This requires addressing the cortisol-dopamine relationship directly: recalibrating the stress response so that oxytocin and dopamine pathways can register social connection as rewarding again. This work is not behavioral. It is neurological. The target is the nervous system’s capacity to receive connection, not just the individual’s willingness to seek it.
The third phase addresses the cognitive patterns that have consolidated around this state: the meaning-making system that has developed to explain and justify the disconnection. This is where the belief that “I am fundamentally different from other people” or “genuine connection is not possible for someone like me” has taken up structural residence in the neural architecture. These are not attitudes. They are learned patterns with neural substrate: patterns often rooted in early attachment styles and relational conditioning. They are changed through the same mechanism that changes any deeply wired pattern: sustained, strategic engagement of neuroplasticity.
The disconnection high-capacity individuals experience is a neurological problem: one that operates across the full domain of relationship intelligence and interpersonal neuroscience. It has a neurological mechanism. It has a neurological solution. The gap between those who resolve it and those who carry it for decades is not a gap in intelligence, insight, or effort: it is a gap in the level of intervention. Those who resolve it work at the level of the nervous system. Those who don’t are working at the level of behavior, which is the wrong address.
If the pattern described here maps onto your experience (the chronic disconnection despite a full life, the fatigue that doesn’t resolve, the hypervigilance in social contexts, the sense that connection is happening around you but not to you) the next step is a conversation. Not to be diagnosed or assessed, but to understand specifically what is running in your neural architecture and what an intervention at that level would look like. schedule a strategy call with Dr. Ceruto.