Attachment as Neural Architecture, Not Personality Type
The conventional framing of attachment styles (secure, anxious, avoidant, disorganized) has become so culturally embedded that most people treat these categories as fixed personality traits. In my practice, I see the consequences of that framing daily: individuals who have internalized their attachment pattern as an identity rather than understanding it as a neural configuration that was built, reinforced, and can be restructured. The neuroscience tells a fundamentally different story than the pop psychology version.
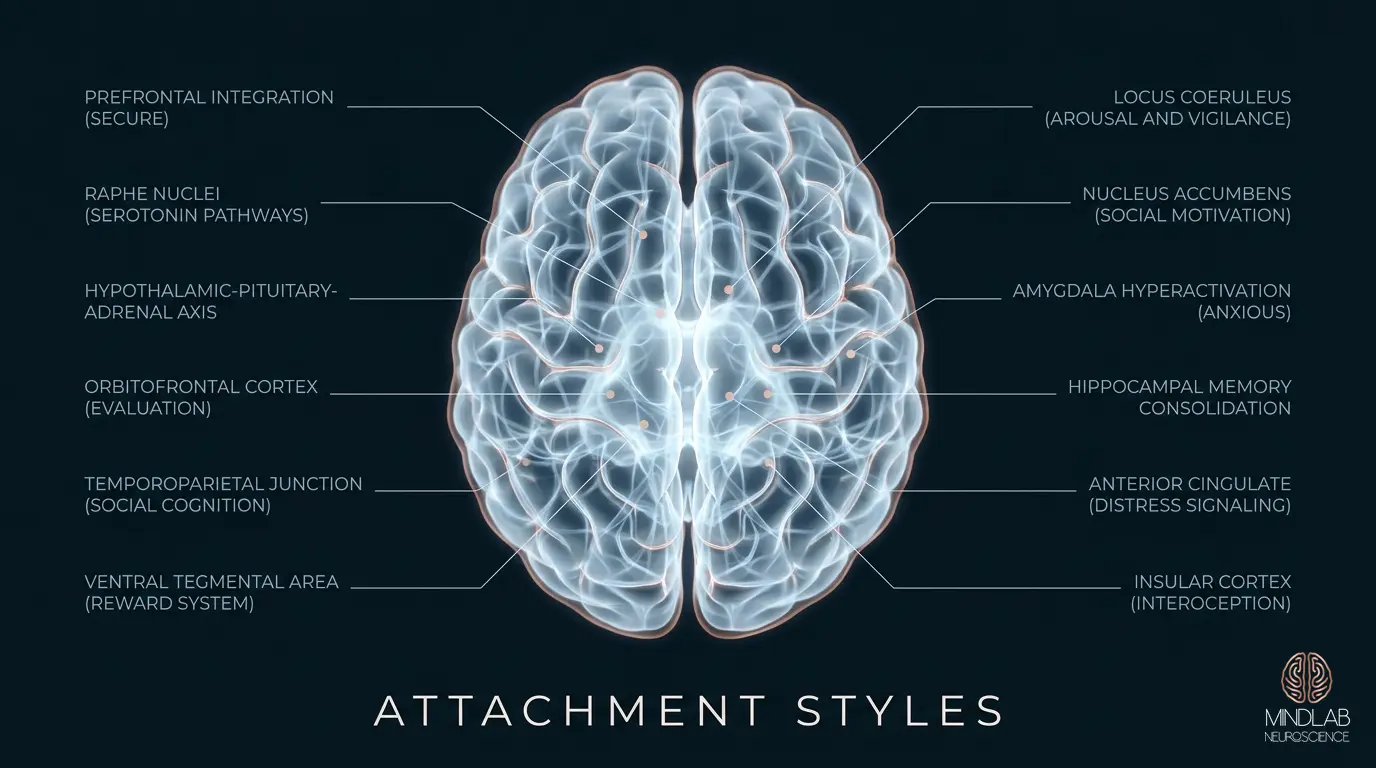
Attachment patterns are not personality types. They are learned neural architectures: specific configurations of connectivity between the prefrontal cortex, the limbic system, and the autonomic nervous system that were shaped during critical developmental windows and subsequently reinforced through thousands of relational interactions. John Bowlby’s original attachment theory, developed in the 1950s, described behavioral patterns. What neuroscience has since revealed is that those behavioral patterns are the surface expression of deeply wired circuitry: circuitry that governs threat detection, reward processing, stress regulation, and the capacity for interpersonal trust.
This distinction matters enormously. When attachment is understood as a personality trait, change feels impossible: you are what you are. When attachment is understood as a neural circuit configuration, change becomes a question of neuroscience: which circuits, which connections, and what conditions allow restructuring? That question has answers. The brain that built one configuration can build another, provided the right conditions are met.
The Oxytocin-Vasopressin Bonding Circuit
At the neurochemical level, attachment is orchestrated by two neuropeptides that operate as a bonding system: oxytocin and vasopressin. These are not the simplistic “love hormones” of popular science articles. They are sophisticated modulators of social cognition, threat appraisal, and reward processing, and the way they function in any given individual is shaped by the attachment architecture that was established in early life.
Oxytocin modulates the amygdala’s threat response to social stimuli, shifting the brain’s default interpretation of ambiguous social signals from threatening toward safe. In securely attached individuals, oxytocin release during close relational contact produces a measurable reduction in amygdala reactivity: the neurochemical signature of interpersonal trust. In individuals with anxious attachment patterns, oxytocin can paradoxically increase vigilance, because the same neurochemical is operating within a circuit that has been calibrated to expect relational threat even during proximity.
Vasopressin plays a complementary role, particularly in bonding behaviors associated with partner selection and pair maintenance. Research from Larry Young’s laboratory at Emory University demonstrated that vasopressin receptor density in the ventral pallidum, a key reward circuit structure, directly predicts bonding behavior. Higher receptor density corresponds to stronger pair-bond formation. Lower density corresponds to reduced bonding motivation. These receptor densities are not immutable. They are shaped by early relational experience and remain modifiable through sustained relational input that meets specific neurochemical conditions.
What I consistently observe in my work is that individuals who describe themselves as “unable to bond” or “incapable of deep connection” are not describing a permanent neurological deficit. They are describing an oxytocin-vasopressin system that was calibrated under conditions of relational unpredictability, and that system can be recalibrated when the right neurological conditions are established.
How Early Caregiving Shapes Prefrontal-Limbic Connectivity
The attachment architecture an individual carries into adulthood was built during a specific developmental window, roughly the first three years of life, when the brain’s prefrontal-limbic connectivity is being established at extraordinary speed. The quality of caregiving during this window literally determines the structural and functional connections between the prefrontal cortex and the limbic system that will govern emotional regulation, threat assessment, and relational behavior for decades.
Consistent, responsive caregiving produces robust connectivity between the medial prefrontal cortex (mPFC) and the amygdala. This connectivity allows the prefrontal cortex to modulate limbic threat signals efficiently: the neural substrate of what we observe behaviorally as emotional regulation and secure attachment. The caregiver functions as an external regulator during the period before the infant’s own prefrontal cortex is sufficiently developed to perform this function independently. Each successful co-regulation episode strengthens the prefrontal-limbic pathway, building the infrastructure the child will eventually use to self-regulate.
Inconsistent caregiving (responsive sometimes, unavailable other times) produces a different architecture. The prefrontal-limbic connections develop, but they are less reliable. The amygdala maintains elevated baseline reactivity because the modulation signal from the prefrontal cortex is intermittent. The result, at the level of neural circuitry, is a brain that is constantly monitoring relational signals for threat or withdrawal: the architecture underlying anxious attachment.
Consistently unavailable or rejecting caregiving produces yet another configuration. The prefrontal cortex develops strong inhibitory connections that suppress limbic activation during relational proximity, not because the emotional response isn’t happening, but because the brain learned early that expressing attachment needs produced negative outcomes. The emotions are present. The neural suppression prevents their expression. This is the architecture that drives avoidant attachment, and understanding it at this level changes everything about how restructuring proceeds.
Earned Secure Attachment Is Neuroplasticity in Action
One of the most significant findings in attachment neuroscience is the concept of earned secure attachment: the empirically validated observation that individuals who began life with insecure attachment patterns can develop secure attachment functioning in adulthood. This is not a theoretical possibility. It is documented in longitudinal research, observable in brain imaging, and something I have witnessed across 26 years of practice with remarkable consistency.
Earned security is neuroplasticity in its most meaningful application. The prefrontal-limbic connectivity patterns that were established in early development are not permanent hardware installations. They are neural pathways that were strengthened through repetition and can be restructured through new relational experiences that meet specific neurological conditions. The key conditions are sustained emotional safety, consistent responsiveness, and, critically, real-time access to the moments when the old attachment pattern activates.
Research from Ruth Feldman’s laboratory has demonstrated that adults who achieve earned security show oxytocin profiles and neural activation patterns that are functionally indistinguishable from individuals who were securely attached from infancy. The brain did not simply compensate. It rebuilt. The prefrontal-limbic connections that were inadequately established in early life were constructed later, through relational experiences that provided the co-regulation the original caregiving environment did not.
This is why I approach attachment work not as a process of managing an insecure style, but as a process of building the neural architecture that the original environment failed to construct. The brain’s plasticity window for attachment-related circuitry does not close after childhood. It narrows, and the conditions required for restructuring become more specific, but the capacity remains. Understanding the neuroscience of the broader neuroscience of relationships makes clear that relational patterns are neural patterns, and neural patterns can be revised.
The Amygdala-Based Threat Detection System in Anxious Attachment
Anxious attachment, viewed through a neuroscience lens, is fundamentally a disruption of threat calibration. The amygdala, specifically the basolateral amygdala, has been tuned to detect relational threat at a lower threshold than the actual environment warrants. Ordinary signals (a delayed text response, a shift in vocal tone, a partner’s momentary distraction) trigger the same neural alarm that would be appropriate for genuine abandonment. The threat detection system is functional. Its calibration is not.
Neuroimaging studies consistently show that individuals with anxious attachment display heightened amygdala reactivity to facial expressions that are ambiguous or mildly negative. Where a securely attached individual’s brain reads a neutral face as neutral, the anxiously attached brain reads it as potentially rejecting. This is not a cognitive distortion in the traditional sense: it is a calibration bias built into the perceptual system itself, operating below the level of conscious interpretation.
The hypothalamic-pituitary-adrenal axis compounds this pattern. In anxious attachment, the HPA axis maintains elevated cortisol baseline levels during relational ambiguity, producing the physiological arousal (the racing heart, the gut tension, the inability to focus) that characterizes attachment anxiety. The individual is not “overreacting.” Their nervous system is responding accurately to the threat signal their amygdala is producing. The problem is upstream: the amygdala’s threat threshold, not the body’s response to it.
What I have observed consistently is that addressing anxious attachment at the behavioral level, teaching someone to “self-soothe” or “communicate their needs”, produces limited durability because it leaves the amygdala calibration unchanged. The behavioral strategies work when consciously deployed, but the moment cognitive resources are depleted (under stress, during conflict, late at night) the amygdala’s original calibration reasserts itself. Lasting change requires recalibrating the threat detection system itself, not layering management strategies on top of it. The connection between attachment anxiety and the body’s broader stress architecture is mapped extensively within emotional resilience and regulation research.
Avoidant Attachment as a Dopaminergic Suppression Pattern
Avoidant attachment presents as emotional distance, self-sufficiency, and discomfort with intimacy. At the neural level, what is actually occurring is a systematic suppression of the brain’s reward circuitry during relational proximity: a pattern that traces directly to dopaminergic pathway modulation established in early development.
In a securely attached brain, close relational contact activates the ventral tegmental area (VTA) and the nucleus accumbens: the core structures of the mesolimbic reward pathway. Proximity to an attachment figure produces dopamine release, which the brain registers as rewarding. This is the neurochemical basis of the felt sense of connection: the brain literally rewards closeness.
In avoidant attachment, this reward signal is attenuated. The VTA-nucleus accumbens circuit shows reduced activation during relational proximity, while simultaneously the dorsolateral prefrontal cortex, the seat of cognitive control, shows increased activation. The brain is actively suppressing the reward response to closeness. This is not a deficit in the capacity to feel. It is a learned suppression pattern: the brain discovered early that attachment-seeking behavior produced pain rather than reward, and it adapted by dampening the very circuit that motivates connection.
The physiological signature is revealing. Individuals with avoidant attachment often report feeling “nothing” during intimate moments, but their autonomic nervous system tells a different story: elevated skin conductance, increased heart rate variability disruption, and cortisol patterns consistent with sympathetic activation. The body is responding. The prefrontal suppression is preventing the conscious experience of that response. The emotions are not absent. They are neurologically muted.
In 26 years of practice, I have found that avoidant attachment is among the most misunderstood patterns, both by the individuals who carry it and by the partners who experience its effects. The avoidant individual is not cold, selfish, or incapable of love. They are operating with a reward circuit that was trained to treat closeness as a threat signal rather than a safety signal. Restructuring this pattern requires retraining the dopaminergic response to relational proximity: rebuilding the reward association that the original environment failed to establish or actively punished.
Restructuring Attachment Circuitry at the Neural Level
Dr. Sydney Ceruto’s approach to attachment work begins from a premise that distinguishes it from virtually every other available option: attachment patterns are neural circuit configurations, and restructuring them requires intervening at the neural level, not the behavioral level, not the cognitive level, and not through insight alone.
The process starts with precision mapping. Which circuits are driving the pattern? Is the primary driver amygdala threat calibration (anxious pattern), dopaminergic reward suppression (avoidant pattern), or the oscillation between both that characterizes disorganized attachment? The answer determines the intervention architecture. A recalibration strategy designed for anxious attachment applied to an avoidant pattern would target the wrong neural substrate, and this category of error is exactly why generic approaches to attachment produce inconsistent results.
Real-Time Neuroplasticity™ addresses attachment circuitry during the moments when those circuits are active: during relational interactions, during conflict, during the specific situations that trigger the pattern. These moments represent heightened plasticity windows, periods when the relevant neural pathways are most available for restructuring. The neuroscience of memory reconsolidation confirms that active neural patterns, when reactivated under specific conditions, can be modified in ways that update the original encoding rather than merely layering new learning on top of it.
This is a critical distinction. Most approaches to attachment work create new learning that competes with the old pattern: a cognitive override that requires conscious effort and tends to fail under stress. Memory reconsolidation, accessed through real-time intervention during pattern activation, modifies the original circuit. The old pattern does not need to be constantly overridden because the circuit itself has been updated.
The individuals I work with often arrive having spent years understanding their attachment pattern intellectually: reading about it, discussing it, identifying it in their relationships. That understanding is not without value, but it has not changed the pattern because insight does not restructure neural circuits. Neural restructuring requires the right conditions: activation of the target circuit, introduction of prediction error (an experience that violates the pattern’s expectation), and consolidation of the new encoding. These conditions are achievable. They are what my methodology is designed to create.
If your attachment patterns are shaping your relationships, your decisions, and your capacity for genuine connection in ways that insight alone has not changed, the next step is a conversation about what recalibration would look like for your specific neural architecture. Schedule a strategy call with Dr. Ceruto to map the circuitry that is driving the pattern and identify what restructuring requires.
What the Neuroscience of Attachment Means for Lasting Change
The neuroscience of attachment has moved decisively beyond the Bowlby-Ainsworth typology that still dominates popular understanding. We now know that attachment patterns are neural circuit configurations involving specific prefrontal-limbic connectivity patterns, specific neurochemical profiles, and specific autonomic nervous system calibrations, all of which were shaped by early relational experience and all of which remain modifiable through the brain’s inherent neuroplasticity.
The oxytocin-vasopressin bonding system, the prefrontal-limbic connectivity architecture, the amygdala’s threat calibration threshold, the dopaminergic reward response to relational proximity: these are not abstract concepts. They are the specific neural substrates that produce the behavioral patterns people experience as their “attachment style.” And each one has a documented pathway to restructuring, provided the conditions are met.
What those conditions require (real-time access to the active circuit, precision identification of the driving substrate, and intervention during heightened plasticity windows) is what distinguishes neuroscience-based attachment work from the approaches that have left so many individuals feeling that their pattern is permanent. The pattern is not permanent. The brain that built it can rebuild it. The question is whether the conditions for that rebuilding are present, and the science of what those conditions are has never been clearer.
Every article in this collection explores attachment through the lens of neural architecture: how the brain builds relational patterns, how those patterns express themselves in behavior, and what the neuroscience reveals about the genuine capacity for change. For deeper exploration of the neural mechanisms underlying trust issues and relational repair, continue through the articles below.