Key Takeaways
- Oxytocin does not generate love. It selectively amplifies trust and social salience toward specific individuals, making a bonded partner feel categorically different from everyone else, regardless of whether the relationship is healthy
- The dopamine surge of early romantic attraction activates the same ventral tegmental area and nucleus accumbens pathways as reward-seeking behavior, the brain classifies new love as a motivation state, not simply an emotion
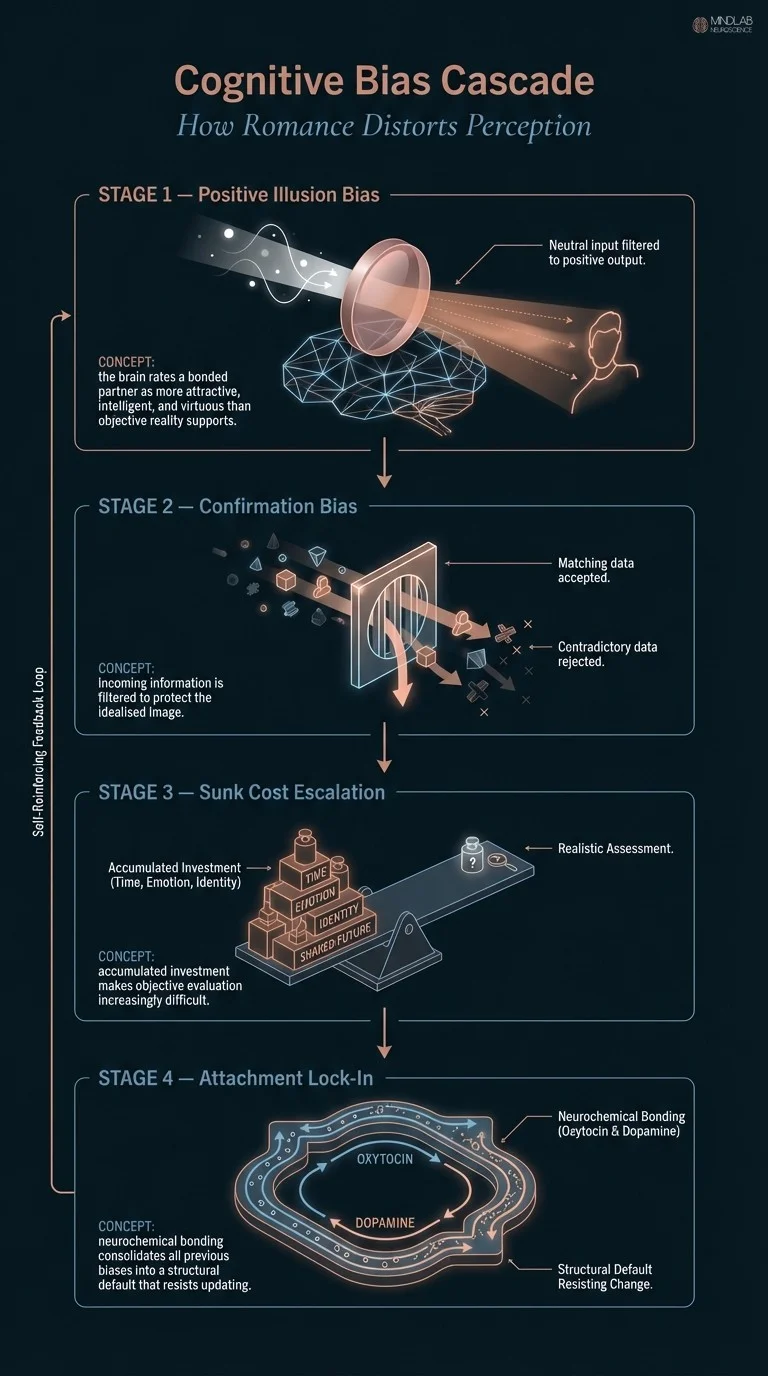
- Cognitive biases in romantic relationships are not failures of intelligence, the brain’s reward and attachment circuitry actively suppresses critical evaluation of a bonded partner, making objective perception neurologically effortful
- Attachment styles are learned neural architectures established through thousands of early caregiver interactions, encoded as default prediction models the brain applies automatically to every subsequent close relationship
- The brain’s love biases can be rewired through deliberate practice that targets automatic appraisal systems during emotionally activated states, not through insight or intellectual understanding alone
The brain does not fall in love rationally. It falls in love through a cascade of neurochemical events, dopamine, oxytocin, norepinephrine, that reshape perception, override critical evaluation, and produce a motivational state powerful enough to reorganize priorities, tolerate significant sacrifice, and sustain pursuit behaviors that would be inexplicable in any other context. At MindLAB Neuroscience, I have spent 26 years working with individuals whose romantic decisions have confounded their own intelligence, choosing partners who replicate harmful patterns, sustaining bonds that produce more distress than connection, or struggling to form authentic intimacy and secure attachment despite deeply wanting to. The neuroscience of love biases explains why these patterns persist and what it takes to restructure them.

What Does Oxytocin Actually Do in Romantic Relationships?
Oxytocin is not simply a “love hormone”. It is a neurochemical that selectively amplifies trust and social salience toward specific individuals, making a bonded partner feel categorically different from everyone else in your world. When oxytocin is released during physical closeness, prolonged eye contact, or shared vulnerability, it does not generate love from nothing, it deepens the neural encoding of that particular person as safe, significant, and irreplaceable, one of the core mechanisms behind the neuroscience of relationship intelligence.
This matters because many individuals arrive confused about why they feel so intensely bonded to someone who may not be treating them well. The oxytocin system does not evaluate merit, it responds to proximity and repetition. The more time two people spend together in emotionally activated states, the more the brain labels that person as central to its survival architecture.
Rigney, de Vries, Petrulis, and Young detailed oxytocin and vasopressin pathways from neural circuits to clinical opportunities, establishing the complexity of these neuropeptide systems in social behavior far beyond the simplified popular narrative. In my practice, I consistently observe that oxytocin-driven bonding forms around distressing relationships just as readily as healthy ones. Shared stress and conflict activate the same release pathways as warmth and affection, which helps explain why volatile relationships can feel paradoxically intense.
The catch is that the neurochemical bond does not carry information about the relationship’s health. It carries information about the relationship’s salience to the nervous system, and those are very different things.
How Does Dopamine Create the Feeling of Falling in Love?
The intoxicating rush of early romantic love is driven largely by dopamine surging through the brain’s reward circuitry, specifically the ventral tegmental area and nucleus accumbens, the same pathways activated by novelty, unpredictability, and reward anticipation. The brain classifies new love as a motivation state, not simply an emotion, which explains its capacity to override sleep, appetite, and prior commitments.
What makes this clinically significant is the mechanism of variable reinforcement. When a new partner is sometimes warm and sometimes distant, dopamine output actually increases. The brain escalates pursuit behavior in response to inconsistency, not consistency. In my practice, I consistently observe that individuals who describe the most “electric” early connections are often describing a dopamine system in overdrive, not compatibility.
Fisher, Aron, and Brown used fMRI to demonstrate that newly love-struck individuals displayed caudate nucleus activation patterns nearly identical to those seen in goal-directed reward-seeking behavior. A motivation state compels action and tolerates significant cost to achieve its objective, which is why early love can override judgment with startling ease.
The dopamine spike of early attraction is temporary by neurological design. When individuals mistake the fading of that spike for the fading of love itself, they make decisions based on a misreading of their own brain chemistry that can dismantle otherwise viable relationships.
The oxytocin system does not evaluate whether a relationship is healthy. It evaluates salience, and those are very different things.
Why Do Cognitive Biases Distort How You See Your Partner?
Romantic relationships are uniquely fertile ground for cognitive distortion because the brain’s reward system and attachment circuitry actively suppress critical evaluation of a bonded partner, making objective perception of that person neurologically effortful. You are not naive when you fail to see your partner clearly, your brain is functioning exactly as it evolved to function.
Several biases cluster reliably in romantic contexts. Positive illusion bias leads individuals to rate their partners as more attractive, more intelligent, and more virtuous than outside observers do. Confirmation bias filters incoming information to protect that idealized image. When a partner does something inconsiderate, the brain hunts for an explanation that preserves the established model rather than updating it.
In 26 years of practice I have found that the individuals who struggle most with this are not lacking in intelligence. They are often among the most cognitively capable people I work with. The bias operates below the level of deliberate reasoning. Rewiring it requires targeted work at the level of automatic appraisal, not simply being told to see things more clearly. The nuance is that some degree of positive bias appears adaptive, relationships require sustained investment, and mildly idealized views sustain motivation through difficulty. The problem arises when idealization becomes so rigid that genuine patterns of harm cannot register at all.
What Role Does the Amygdala Play in Romantic Attraction and Mate Selection?
The amygdala functions as a rapid threat-and-reward evaluator, processing a potential partner’s face, voice, and movement within milliseconds and generating a felt sense of safety or danger before conscious evaluation begins. That immediate gut reaction upon meeting someone is substantially an amygdala output.
This matters for understanding why attraction so often defies rational explanation. The amygdala cross-references incoming sensory data against a lifetime of stored emotional memory, including early attachment experiences with caregivers. A stranger’s vocal tone or interpersonal energy may activate patterns encoded decades earlier, producing attraction or aversion that feels inexplicable but is deeply patterned.
Shih and colleagues conducted a meta-analysis of neuroimaging studies of maternal and passionate love, confirming that both activate overlapping reward and attachment regions while showing distinct regulatory patterns in the amygdala and social cognition networks. In my practice, I find that an individual who grew up in an unpredictable household may experience a calm, reliable partner as flat or unexciting, while feeling a charged pull toward someone whose relational style mirrors the chaos they were conditioned to navigate.
Recognizing the amygdala’s role does not neutralize these reactions, but it creates the cognitive distance necessary to evaluate them rather than act on them automatically.
Does Romantic Love Really Work Like an Addiction in the Brain?
At a neurochemical level, intense romantic attachment activates the same dopaminergic reward pathways, produces the same craving-and-withdrawal dynamics, and shows the same tolerance escalation as substance dependence, which is not a metaphor but a structural overlap confirmed by neuroimaging. The similarity is anatomical.
This framing is useful precisely because it reframes heartbreak. When a bonded relationship ends, the brain enters a neurochemical withdrawal state, dopamine and oxytocin signaling drops sharply, the reward circuitry goes quiet, and the stress response escalates. Individuals describe this as physical pain, and functional imaging confirms that social rejection activates the same anterior cingulate cortex regions as physical pain stimuli.
Fisher, Aron, and Brown established that romantic love functions as a mammalian brain system for mate choice, with neurological infrastructure that serves both attachment and addiction because attachment itself is a survival-critical behavior. The clinical relevance is that attempting to end a harmful attachment through willpower alone is asking the prefrontal cortex to override a system classified as survival-relevant, an extraordinarily difficult ask without structured neural restructuring of romantic attachment circuits.
The critical nuance: this overlap does not mean love is pathological. The same infrastructure serves both because the brain treats bonded attachment with the urgency and priority it applies to food or water.
Attempting to end a harmful attachment through willpower alone is asking the prefrontal cortex to override a system the brain has classified as survival-relevant, an extraordinarily difficult ask without structured neural support.
How Do Attachment Styles Become Wired into Neural Circuitry?
Attachment styles are not personality traits or emotional preferences. They are learned neural architectures, established through thousands of repetitions of early caregiver interactions and encoded as default prediction models that the brain applies automatically to every subsequent close relationship. They persist not because people choose them but because they are structurally embedded.
The brain’s default mode network, which activates during social cognition and self-referential thought, carries these relational models forward through time. When you enter a new relationship, your brain runs its existing model, secure, anxious, avoidant, or disorganized, and generates expectations, emotional reactions, and behavioral impulses accordingly, often before conscious deliberation engages.
Love established that oxytocin interacts with dopamine systems to modulate motivation and social bonding, demonstrating that attachment is not a single-neurotransmitter phenomenon but a multi-system neural architecture involving reward, motivation, and threat-evaluation circuits working in coordination. In my practice, understanding that attachment style is architecture rather than choice transforms how individuals approach the work, from self-blame for their relational patterns to structured restructuring of the circuits producing them.
The important qualification: wired does not mean fixed. The same plasticity that allowed early experiences to shape these systems remains available throughout adulthood, though it requires more deliberate effort. Neural pathways reinforced over decades have significant inertia, rewriting them is not an intellectual exercise but a repetitive experiential one.
How Does the Brain Distinguish Between Genuine Connection and Neurochemical Intensity?
The brain does not naturally distinguish between genuine relational connection and neurochemical intensity, the prefrontal cortex must learn to evaluate what the limbic system is presenting as compelling, and that evaluative capacity is a skill that must be built deliberately rather than one that develops automatically with romantic experience.
Early romantic attraction generates signals, dopamine-driven motivation, oxytocin-mediated bonding, norepinephrine-fueled arousal, that the brain interprets as evidence of exceptional compatibility. But these signals are responses to neurochemical conditions, not assessments of relational health. A volatile dynamic produces more neurochemical activation than a stable one. An intermittently available partner generates more dopamine than a consistently present one. The brain’s reward system equates intensity with value, and this equation produces predictable errors in partner evaluation.
In my practice, the intervention that produces the most reliable shift is building the prefrontal cortex’s capacity to hold the neurochemical experience at a slight evaluative distance, experiencing the pull of attraction without automatically acting on its narrative about what the attraction means. This is not suppression, which would be neurologically counterproductive. It is the development of a parallel processing channel: one track experiencing the neurochemistry, another track evaluating whether the neurochemistry is tracking something genuinely good or something merely familiar. That dual-track capacity is the neural substrate of wise partner selection, and it does not develop through heartbreak alone, it requires structured methodology.
Can You Rewire the Brain’s Love Biases Through Deliberate Practice?
The brain’s love biases can be rewired through neuroplasticity operating through a specific pathway: new relational experience, consistently repeated in emotionally activated states, gradually overwrites maladaptive prediction models at the level of synaptic structure. This is not positive thinking. It is how the brain physically changes.
The prefrontal cortex has direct inhibitory connections to the amygdala and reward circuitry. When an individual repeatedly practices noticing automatic romantic reactions, the pull toward familiar-but-harmful patterns, the dismissal of stable-but-unfamiliar partners, and holds those reactions under reflective examination rather than acting on them, the prefrontal-to-amygdala pathway strengthens. Over time, the automatic reaction loses some of its force.
Seshadri reviewed the neuroendocrinology of love, demonstrating that the hormonal and neural systems underlying romantic attachment are responsive to experiential input across the lifespan, not solely during developmental windows. At MindLAB, the methodology targets change at the level of automatic appraisal and embodied response, not conceptual understanding. What distinguishes lasting change from temporary behavioral adjustment is whether the work reaches deep enough to alter the prediction model itself, not just the surface behavior it produces. The individuals I work with who achieve the most durable transformations in their capacity for genuine intimacy and vulnerability are those who engage with the restructuring process during live relational moments rather than analyzing their patterns retrospectively.
Frequently Asked Questions
How long does the brain’s infatuation phase last?
The dopamine-driven infatuation phase typically lasts between six months and two years before the brain’s reward system habituates to the partner’s presence and dopamine output returns to baseline levels. This neurochemical transition is normal and universal; it reflects the brain shifting from novelty-driven motivation to attachment-based bonding mediated primarily by oxytocin and vasopressin systems. The transition does not indicate diminished love but a change in its neurological substrate, moving from an intense motivational state to a calmer bonding state that is better suited for sustained partnership.
Can you change your attachment style as an adult?
Attachment style can be restructured in adulthood because the neural circuitry underlying attachment retains significant plasticity throughout the lifespan, though the process requires more deliberate effort than the effortless encoding that occurred during early development. Earned secure attachment, security developed through adult relational experiences rather than childhood origins, is well documented in attachment research and produces neural signatures that overlap substantially with developmental security. The restructuring requires consistent new relational experiences that disconfirm the existing prediction model, repeated enough times to overwrite the default architecture at the synaptic level.
Why is heartbreak so physically painful?
Heartbreak produces physical pain because social rejection activates the same anterior cingulate cortex regions and neural pain pathways as physical injury, a finding confirmed by multiple neuroimaging studies. The brain processes the loss of a bonded attachment figure as a survival-relevant threat, triggering a neurochemical withdrawal state comparable to substance withdrawal: dopamine and oxytocin signaling drops sharply, cortisol spikes, and the reward circuits that were routinely activated by the partner’s presence go quiet. The physical symptoms (chest tightness, sleep disruption, appetite changes, difficulty concentrating) are genuine neurological events, not metaphorical distress.
Does the brain process romantic love differently from other forms of love?
Romantic love activates a distinct neural signature compared to parental, platonic, or compassionate love, specifically recruiting the brain’s dopaminergic motivation circuitry, the ventral tegmental area, and the caudate nucleus in a pattern unique to romantic attachment. Parental love shares some overlapping regions but engages additional circuits related to caregiving and threat vigilance. Friendship and compassionate love activate empathy and social cognition networks with less involvement of the high-intensity reward pathways. Romantic love is neurologically distinguished by its motivational intensity and its capacity to produce both the euphoria of reciprocated attachment and the withdrawal-like distress of its loss.
References
- Feldman, R. (2012). Oxytocin and social affiliation in humans. Hormones and Behavior, 61(3), 380-391. https://doi.org/10.1016/j.yhbeh.2012.01.008
- Shamay-Tsoory, S. G., and Abu-Akel, A. (2016). The social salience hypothesis of oxytocin. Biological Psychiatry, 79(3), 194-202. https://doi.org/10.1016/j.biopsych.2015.07.020
- Eisenberger, N. I. (2012). The pain of social disconnection: Examining the shared neural underpinnings of physical and social pain. Nature Reviews Neuroscience, 13(6), 421-434. https://doi.org/10.1038/nrn3231
- Coan, J. A., Schaefer, H. S., and Davidson, R. J. (2006). Lending a hand: Social regulation of the neural response to threat. Psychological Science, 17(12), 1032-1039. https://doi.org/10.1111/j.1467-9280.2006.01832.x
- Kahneman, D., and Tversky, A. (1979). Prospect theory: An analysis of decision under risk. Econometrica, 47(2), 263-292. https://doi.org/10.2307/1914185
- Beck, A. T. (1976). Cognitive therapy and the emotional disorders. International Universities Press.
What the First Conversation Looks Like
When someone reaches out to MindLAB Neuroscience about relationship patterns, they often describe the same frustrating cycle, falling into the same dynamics, choosing partners who trigger the same responses, or losing clarity about their own needs once emotional bonding activates. The patterns feel involuntary because they are: the brain is running prediction models built from earlier relational experience.
Dr. Ceruto will map the neural circuitry driving your specific relational patterns, identifying where oxytocin bonding systems, dopaminergic reward loops, and attachment prediction models are generating the responses you want to change. From that assessment, a structured intervention becomes clear: precise, grounded in how the brain actually forms and revises attachment architecture, and designed to produce measurable shifts in how you experience connection.