Key Takeaways
- Neuroplasticity is the brain’s intrinsic capacity to reorganize its structure and function in response to experience, forming, strengthening, and eliminating synaptic connections across every stage of the lifespan
- Long-term potentiation, the cellular mechanism through which repeated neural activation strengthens synaptic connections, is the principal biological process converting learning into lasting memory
- Synaptic pruning eliminates weak or underused connections during development, increasing processing speed and cognitive efficiency through specialization rather than complexity
- Myelination increases neural signal transmission speed by up to one hundredfold and continues into the mid-twenties, which is why executive function and impulse regulation mature relatively late in development
- Critical periods are genetically timed windows of heightened plasticity, but adult neuroplasticity remains available through experience-dependent mechanisms that require greater deliberate effort and consistency
The human brain is not a fixed organ that finishes developing and then degrades. It is a continuously adaptive system that rewires itself in response to every meaningful experience across the entire lifespan. At MindLAB Neuroscience, this principle is the foundation of every methodology I use. When individuals arrive convinced that their cognitive patterns, emotional responses, or behavioral defaults are permanent features of who they are, the neuroscience of brain plasticity and neural adaptation tells a fundamentally different story. The same mechanisms that built the current architecture can be harnessed to restructure it, with the right methodology, sufficient repetition, and the neurochemical conditions that signal biological relevance to the learning system.
What Is Neuroplasticity and How Does It Shape Brain Development?
Neuroplasticity is the brain’s intrinsic capacity to reorganize its structure and function (forming new synaptic connections, pruning underused ones, and remapping functional territories) in response to experience, learning, injury, and intentional practice, across every stage of the lifespan from prenatal development through advanced age.
Brain development is not a process that happens to a passive organ. It is a continuous dialogue between genetic blueprint and lived experience. From the moment neural activity begins in the developing embryo, the brain is selecting, strengthening, and eliminating connections based on what it encounters. The connections that are activated repeatedly are preserved and fortified; those that go unused are systematically dismantled.
Pascual-Leone and colleagues provided compelling evidence that even mental rehearsal, imagining a skill without physically performing it, produces measurable cortical reorganization, demonstrating that experience-dependent plasticity is driven by neural activation patterns, not solely by physical action. In my practice, this finding has direct implications: the brain does not require a changed external circumstance to begin changing. It requires changed patterns of activation.
The caveat that grounds all of this: plasticity is not infinite or unconditional. It operates within biological constraints that shift across developmental stages, and it is always in competition with the existing architecture, which has the advantage of being already wired, rehearsed, and consolidated.
How Does Long-Term Potentiation Drive Learning and Memory Formation?
Long-term potentiation is the cellular mechanism through which repeated neural activation strengthens synaptic connections (making the firing of a given neural circuit progressively faster, more reliable, and more automatic) and it is the principal biological process by which learning is converted into lasting memory.
When two neurons fire in close temporal sequence, the synapse between them is physically modified. The postsynaptic receptor density increases, the connection’s efficiency improves, and the threshold for future activation is lowered. This is a measurable molecular event involving glutamate receptors, calcium ion cascades, and structural protein synthesis at the synapse.
Lynch provided a comprehensive review of long-term potentiation and its relationship to memory formation, establishing the experimental foundation for understanding how repetition produces synaptic change. In my practice, this mechanism is the neurological justification for why consistency matters more than intensity in any meaningful change process. A single powerful insight does not produce lasting neural change. Repeated activation of the new pattern across multiple contexts is what converts experience into architecture.
What is often misunderstood is the specificity of long-term potentiation: it strengthens exactly the circuit that is activated, not some generalized version of it. Practicing a skill under conditions that differ substantially from its real-world application will build a circuit that does not transfer cleanly.
The brain does not require a changed external circumstance to begin changing. It requires changed patterns of activation, and the restructuring begins at the cellular level with each repetition.
What Happens During Synaptic Pruning and Why Is It Essential?
Synaptic pruning is the brain’s genetically programmed process of eliminating weak, redundant, or underused synaptic connections (reducing neural noise, increasing processing speed, and concentrating resources in the most frequently used circuits) and it is as critical to cognitive development as the formation of new connections.
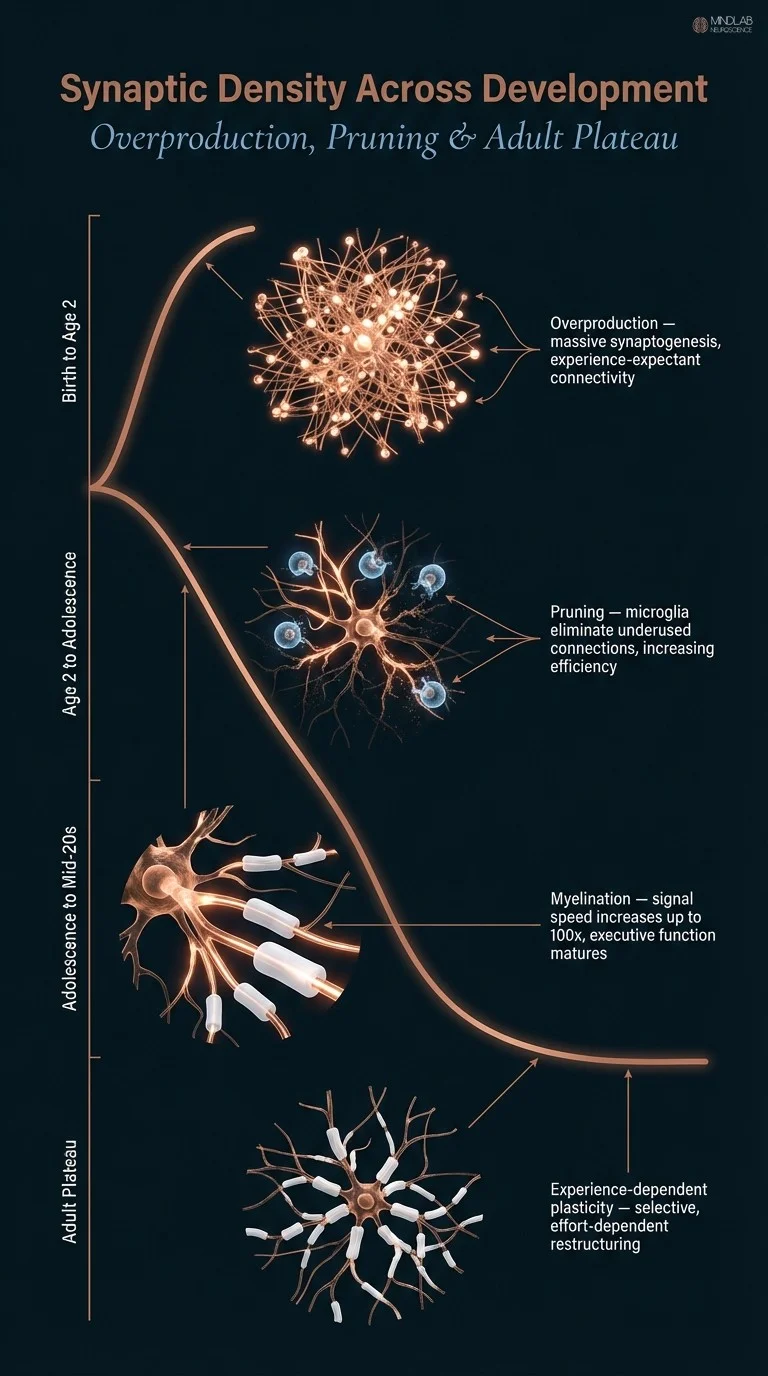
The brain does not grow continuously toward greater complexity. Its development is also a sustained act of subtraction. During infancy, the brain overproduces synaptic connections at a rate that could not be sustained, then systematically eliminates those not being activated by experience. This pruning is not loss. It is specialization.
Paolicelli and colleagues demonstrated that synaptic pruning by microglia is necessary for normal brain development, establishing at the molecular level that the brain’s immune cells actively participate in sculpting neural architecture. In my practice, this process offers a powerful reframe for individuals who feel that developmental difficulties somehow permanently limited them: the pruning that occurred was guided by the experiences available. Different experiences would have guided different pruning.
The nuance is that pruning is not fully reversible. Connections that have been eliminated cannot be simply restored. What can happen (and does, through deliberate, experience-dependent practice) is the formation of new circuits that accomplish similar functions through different architecture.
How Does Myelination Affect Cognitive Speed and Brain Efficiency?
Myelination, the progressive wrapping of axons in a fatty insulating sheath by oligodendrocyte cells, increases neural signal transmission speed by up to one hundredfold, dramatically improving cognitive processing speed, working memory efficiency, and the synchronization of activity across distributed brain networks.
The difference between an unmyelinated axon and a fully myelinated one is qualitative. Myelinated axons transmit signals through saltatory conduction, where the electrical impulse jumps between gaps in the myelin sheath rather than traveling continuously along the membrane. The gradual myelination of the prefrontal cortex, which continues well into the mid-twenties, is the primary neurological reason that executive function, impulse regulation, and long-range planning mature relatively late in development.
Neuroscience research has highlighted the underappreciated role of white matter myelination in complex cognitive functions, demonstrating that myelin development is experience-sensitive and continues to respond to learning and practice in adulthood. In my practice, this has direct relevance for individuals who conclude that their cognitive limitations are permanent because they were present in adolescence. The prefrontal circuitry they are working with now is not identical to what they had at eighteen.
Myelination also explains something individuals often describe as a felt sense: the difference between a skill that requires conscious effort and one that has become automatic reflects, in part, the myelination of the relevant circuits through repeated practice.
What Are Critical Periods in Brain Development, and Do They Ever Close?
Critical periods are genetically timed developmental windows during which the brain is maximally sensitive to specific categories of experience (such as language acquisition, binocular vision, and social attachment) and during which the absence of appropriate input produces structural deficits that become progressively harder to remediate.
The existence of critical periods reflects the brain’s strategy of using early experience to calibrate systems whose optimal configuration cannot be fully specified in advance by genetics alone. The visual cortex requires patterned light input during a specific postnatal window to organize normally. Language acquisition relies on exposure during a period when phonemic discrimination circuits are maximally plastic.
Hensch detailed the molecular mechanisms that open and close critical periods, identifying the role of parvalbumin interneurons and the maturation of GABAergic inhibitory circuits as key regulators. In my practice, critical period research matters not because it defines ceilings but because it establishes the baseline: understanding what was consolidated early informs what restructuring work is realistic and what timelines are truthful.
Critical periods close, but plasticity does not end. What changes is the mechanism and the required investment. Adult plasticity is slower, demands greater effort, and typically requires more deliberate methodology than the effortless acquisition of early development. None of that means it is unavailable.
The pruning that occurred during your development was guided by the experiences available at the time: different experiences would have guided different pruning, and the brain retains the capacity to build new circuits at any age.
How Does Experience-Dependent Plasticity Rewire the Adult Brain?
Experience-dependent plasticity rewires the adult brain through the same Hebbian mechanism that shapes early development, neurons that fire together wire together, but with a higher activation threshold, a slower consolidation rate, and a greater dependence on attention, repetition, and the neurochemical conditions that signal biological relevance to the learning system.
The adult brain does not stop changing. It becomes more selective about what it changes for. Novel, meaningful, emotionally salient, or repeatedly activated experiences still drive synaptic modification in adulthood. The difference from childhood plasticity is one of efficiency: the young brain acquires new patterns with relatively less effort because its plasticity mechanisms are in an open, highly sensitive state.
Kolb and Gibb documented that brain plasticity and behaviour are deeply interlinked across the developing brain, establishing principles that continue to apply, with modified parameters, throughout the adult lifespan. In my practice, this is one of the most useful reframes: the brain you use differently physically becomes different. And the change is detectable on a scanner within months.
What experience-dependent plasticity requires in adulthood, and what most approaches to personal change fail to provide, is sufficient repetition across sufficient time, under conditions of attentional engagement and genuine neurochemical investment. Passive exposure does not drive robust plasticity. Deliberate, consistent, motivated practice does. This is why Real-Time Neuroplasticity™ works during live emotional moments: the brain’s plasticity mechanisms are most engaged when the relevant circuits are actively firing under conditions the brain registers as genuinely important. This connects directly to the neural drivers of to rewire your brain.
Can Stress and Trauma Alter the Brain’s Plasticity Mechanisms?
Chronic stress and unresolved trauma fundamentally alter the brain’s plasticity mechanisms, shifting the default direction of neural change from adaptive learning toward threat-sensitization, reducing hippocampal plasticity while amplifying amygdala reactivity, and creating a neurological state in which the brain is simultaneously less able to learn new patterns and more efficient at reinforcing defensive ones. For how this fits the larger picture, see how the brain calms and re-regulates after stress.
Under normal conditions, plasticity is relatively balanced, the brain strengthens useful circuits and weakens unused ones based on experience. Under chronic stress, cortisol exposure disrupts this balance. The hippocampus, which is dense with cortisol receptors, loses dendritic complexity and reduced capacity for new learning. The amygdala, by contrast, shows enhanced plasticity under stress: its threat-detection circuits become more sensitive and more efficient, which is adaptive in a genuinely dangerous environment but counterproductive when the stress is internal or relational rather than environmental.
In my practice, this explains a pattern I encounter regularly: individuals who are extraordinarily quick to detect threat, criticism, or social exclusion but who struggle to consolidate new, adaptive responses. Their brain’s plasticity has not been lost. It has been redirected. The amygdala is learning rapidly. The prefrontal cortex and hippocampus are learning slowly. Reversing this imbalance requires methodology that specifically targets prefrontal-hippocampal plasticity while gradually recalibrating amygdala sensitivity, work that must happen under conditions of neural safety for the cognitive flexibility circuits to re-engage.
What Is Hebbian Learning and How Does It Shape Neural Circuits?
Hebbian learning, summarized as “neurons that fire together wire together”, is the foundational principle of synaptic modification, describing how the simultaneous activation of pre- and postsynaptic neurons strengthens the connection between them, gradually assembling coordinated neural circuits from individual activation events.
Donald Hebb proposed this principle in 1949, and decades of subsequent molecular neuroscience have confirmed and elaborated the mechanism. When a presynaptic neuron fires and its activation contributes to the firing of a postsynaptic neuron, the synapse between them is tagged for strengthening. Over repeated co-activations, this strengthening becomes structural.
In my practice, Hebbian learning is the neurological justification for every structured repetition built into a methodology: the goal is not to rehearse a behavior but to co-activate the neurons that need to become a circuit. The behavioral repetition is the delivery mechanism for the neural event that actually matters. The individuals who understand this (that they are building architecture, not just practicing habits) approach the work with a precision that accelerates their neuroplasticity-driven personal transformation.
The implication that carries the most weight for adults undertaking deliberate change work is directional and often misunderstood: Hebbian learning is bidirectional. The circuits that are repeatedly co-activated grow stronger through long-term potentiation. The circuits that are repeatedly not co-activated, because a new pattern is being practiced in their place, grow weaker through a complementary process called long-term depression. You are not simply adding new circuits alongside old ones. You are, over time, actively diminishing the synaptic strength of the patterns you are replacing. That bidirectional mechanism is what makes lasting change neurologically possible, and it is why consistency is not optional, every repetition of the new pattern simultaneously builds the replacement and weakens the original.
Frequently Asked Questions
What activities best promote neuroplasticity in adults?
Activities that most effectively promote adult neuroplasticity share three characteristics: novelty, emotional engagement, and progressive challenge that slightly exceeds current capacity. The brain’s plasticity mechanisms are most responsive when an activity requires focused attention, involves genuinely new learning rather than repetition of mastered skills, and carries emotional or motivational significance that signals biological relevance. Physical exercise increases brain-derived neurotrophic factor, which supports synaptic growth and survival. Complex skill acquisition restructures motor and cognitive cortex. Emotionally engaged relational experiences reshape attachment and regulatory circuits.
Can neuroplasticity reverse brain damage?
Neuroplasticity cannot regenerate destroyed brain tissue, but it can recruit adjacent neural regions to assume functions previously handled by damaged areas, a process called cortical remapping that underlies much of the functional recovery observed after stroke, traumatic brain injury, and other neurological insults. The brain compensates for structural damage by reorganizing existing circuits, strengthening alternative pathways, and in some cases generating new connections that bypass the damaged region entirely. The degree of functional recovery depends on the location and extent of damage, the age of the individual, and the quality of structured rehabilitation.
How long does it take for the brain to form new neural pathways?
New neural pathways begin forming within minutes of novel experience through initial synaptic modification, but these early changes are fragile and require consolidation through sleep and repeated activation to become structurally durable. Measurable changes in brain connectivity appear on neuroimaging within four to eight weeks of consistent, focused practice. Full structural consolidation, including protein synthesis, dendritic spine growth, and potential myelination of the new pathway, requires three to six months of sustained engagement. The popular notion of twenty-one days to form a habit has no neurological basis; meaningful neural restructuring operates on a longer timeline.
What is the difference between neuroplasticity and neurogenesis?
Neuroplasticity refers to the brain’s capacity to reorganize existing neural connections, strengthening, weakening, forming, or eliminating synapses between existing neurons in response to experience. Neurogenesis is the generation of entirely new neurons, which in the adult brain occurs primarily in the hippocampus and olfactory bulb. Both processes contribute to the brain’s adaptability, but neuroplasticity is the far more widespread and functionally significant mechanism for adult learning and behavioral change. Most of the brain changes associated with skill acquisition, emotional restructuring, and cognitive development in adulthood are driven by synaptic plasticity rather than neurogenesis.
References
- Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn, U., and May, A. (2004). Neuroplasticity: Changes in grey matter induced by training. Nature, 427(6972), 311-312. https://doi.org/10.1038/427311a
- Pascual-Leone, A., Amedi, A., Fregni, F., and Merabet, L. B. (2005). The plastic human brain cortex. Annual Review of Neuroscience, 28, 377-401. https://doi.org/10.1146/annurev.neuro.27.070203.144216
- Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D., Ashburner, J., Frackowiak, R. S., and Frith, C. D. (2000). Navigation-related structural change in the hippocampi of taxi drivers. Proceedings of the National Academy of Sciences, 97(8), 4398-4403. https://doi.org/10.1073/pnas.070039597
- Kolb, B., and Gibb, R. (2011). Brain plasticity and behaviour in the developing brain. Journal of the Canadian Academy of Child and Adolescent Psychiatry, 20(4), 265-276. https://pubmed.ncbi.nlm.nih.gov/22114608/
- Kandel, E. R., Schwartz, J. H., Jessell, T. M., Siegelbaum, S. A., and Hudspeth, A. J. (2013). Principles of Neural Science, 5th Edition. McGraw-Hill.
- Damasio, A. R. (1996). The somatic marker hypothesis and the possible functions of the prefrontal cortex. Philosophical Transactions of the Royal Society B, 351(1346), 1413-1420. https://doi.org/10.1098/rstb.1996.0125
What the First Conversation Looks Like
When someone reaches out to MindLAB Neuroscience about neuroplasticity, they are usually past the general concept: they understand the brain can change. What they want to know is whether their specific patterns, habits, or cognitive limitations can actually be restructured, and what that process looks like in practice rather than in theory.
Dr. Ceruto will assess the specific neural systems involved in your situation, identifying which circuits have consolidated through repetition, where plasticity is most accessible, and what structured intervention will produce measurable change within your particular neurological profile. From that assessment, a precise path forward becomes visible: targeted to your architecture, grounded in current neuroscience, and designed to produce results you can track.